
6 Paleoanthropology
Phil Geib; Bill Belcher; and Timothy Sefczek
Learning Objectives
|
Human Evolution
It is important to know that the human body in its modern form, including our biology, physiology, morphology, etc., is the result of evolution by natural selection. We are the products of a long evolutionary history. The products of evolution operating over millions of years, slowly shaping us into what we are today. Consequently, much about our behavior can be understood by knowing how natural selection works and knowing what kinds of traits it favored during our evolution.
Our long history of evolution by natural selection began with the first appearance of primates some 65 million years ago, after extinction of the dinosaurs. In the primate lineage the first monkeys appeared around 40 million years ago with apes splitting from monkeys at about 25 million. At about 13 million years ago, the ape-like ancestors of gorillas, chimps & humans appeared. The gorilla line split off about 10 million and the chimp-human line split 6.5-9.3 million. The first bipedal-ancestor appears around 7 million years ago, and an ancestor with a human-like form appears just under 2 million years ago. But fully modern humans didn’t evolve until around 200,000 years ago. These time spans allow for significant change from evolution by natural selection.
It is also important to recognize that human evolution continues. We are still evolving. It has not ended because evolution never ceases. Evolution is not a goal driven process. There is no such thing as a perfectly adapted organism, the pinnacle of a species, because adaptation depends on the environmental context and that is constantly in flux.
There is now a coevolution that occurs between our genes and our culture. This gene-culture coevolution is evident in such traits as lactase persistence—the ability of many adults to produce the enzyme to break down lactose in milk. When this trait appeared in some humans, which it does on occasion by random, it spread because it conferred a reproductive advantage for those who started to drink the milk of newly domesticated livestock. The spread of this trait can be tracked by examining prehistoric DNA. As will be discussed in more detail in the chapter on archaeology, the frozen ancient corpse found on Italian and Austrian boundary known as Ötzi was lactose intolerant, indicating that this trait was perhaps not yet common some 5200 years ago.
Chimps, bonobos, and humans are closely related genetically and African in origin, but many characteristics set us apart from these other two species. It is these characteristics, which are discussed below, that some believe justify making the human lineage distinct. In this text we use the term hominin exclusive of chimpanzees and bonobos, the two Pan species that are often classified as part of the hominin tribe (Hominini). Herein, hominin refers to modern humans (Homo sapiens) and all ancestors back to the split from Pan roughly 6.5-9.3 million years ago.
Distinctive Human Physical Traits
Bipedalism
Humans are the only primates that are obligate bipeds. This was not always the case, even as recent 50,000 years ago there were at least three other bipedal hominins aside from us (Homo floresiensis, Homo luzonensis, and Neanderthals) and further back in time there were more. We have several skeletal adaptations that enhance bipedalism such as the shape of our pelvis, the curvature of our lumbar vertebrae, and our arched, non-prehensile feet. These traits are not seen in our closest evolutionary cousins the chimps and bonobos or any other apes or monkeys.
A series of evolutionary skeletal changes occurred millions of years ago to accommodate bipedalism. Some of these occurred to the foot, some to the leg bones, some to the pelvis, some to the backbone and some to the skull. It was a modification from toes right to up to the connection with the head. Once hominins adapted this mode of locomotion, they became more efficient at it and less efficient at any other locomotor form. Some of these changes relate to shifting the balance of body weight so that it puts less strain on muscles.
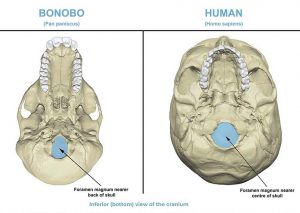
Starting up top, let’s consider the foramen magnum, that hole in the base of your skull through which the spinal cord passes. In fully quadrupedal animals that maintain a horizontal body position most of the time, such as a dog, this essential skeletal feature occurs at the back of the skull, a fitting place given their means of locomotion. For knuckle-walking chimpanzees and bonobos this hole is more underneath the skull (more central), though not as far as humans, as primates tend towards a more upright posture. Both chimpanzees and bonobos move quadrupedally but spend considerable time with their bodies at a backward incline to the world, in part because their arms are longer, but also related to their arboreal adaptation. With bipedal movement, if the foramen magnum was not positioned more forward or centrally on the skull, as shown above, then your head would naturally face upwards, an awkward position for observing the world at ground level.
Next down the body is the spinal connection to the head, especially the thoracic vertebrae (those of the upper body with ribs attached that extend from the neck down to the abdomen) and the lumbar vertebrae (lower back). The spine became curved in both of these areas, resulting in an S-shaped vertebral column rather than a straight one (lumbar lordosis). The thoracic curve is backward, the lumbar one is forward. These curves help to absorb shock when walking and help keep weight centered above the pelvis.
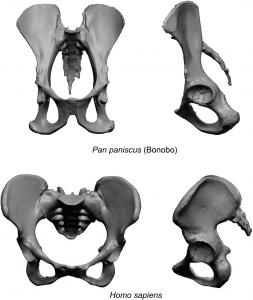
Next descending down the body is the pelvis, which became shorter and broader. The pelvis consists of three bones: two hip bones, or os coxae (pronounced os kok-suh), and the sacrum, which is basically several fused vertebra. In early hominins, the sacrum and hip bones are relatively short compared to apes. The upper portion of each hip bone, called the ilium, is short and curved compared to the long, flattened ilium of chimpanzees and other apes. The curvature places quadricep muscle attachment closer to the front of the body, allowing the muscle greater leverage in pulling the femur forward in an upright posture.
Other muscles of the hip and leg are also modified. The attachment surface of the gluteus maximus is farther in back of the hip joint in humans than chimpanzee. The hamstrings are farther in back of the knee in chimpanzees. For an informative video on comparing human and Pan bipedal walking watch the short video at this link.
Below the hips, structural alterations with bipedalism include limb lengthening and femur angling. Our lower limbs or legs are elongated compared to other living apes. The human thigh bone (femur) comprises approximately 20% of body height, while in gorillas it comprises only about 11%. The calf bone (tibia) comprises 9% of body height in humans but just around 4% in gorillas. The femur in not only longer but angled inward. This keeps the legs and feet more directly under the body. Knee anatomy is also modified to permit full leg extension.
The feet also underwent modifications, which should come as no surprise since they provide the basal support for this whole system. The most distinctive modifications occur to the foot arch and the big toe, but hominin feet also have a robust heel, since this is commonly the first part to touch down in normal walking. There longitudinal arch helps absorb shock and adds a propulsive spring to the step. The big toe is brought in line with the other toes. It no longer diverges like those of all other primates.
One important facet of all these skeletal traits linked to locomotion is that they can get fossilized and thus provide tangible evidence in the paleontological record as to when the traits appeared. This is not true, at least not directly, for some of the other key distinctions of the human lineage such as language; that just does not fossilize very well.
Brains
Our brains are large and complex, much more so than other apes. We have a particularly well-developed cerebral cortex, the place where speech is processed and where other higher mental activities occur. Human brains average about 1300-1400 cc (cubic cementers). In contrast, the massive gorilla has a brain averaging about 525 cc. Gorillas are more massive than us yet our brain is three times larger than their brain. Chimpanzees have a cranial capacity that ranges between 300 and 500 cc but much smaller body mass than gorillas or us, thus they have a larger brain-to-body size ratio than gorillas, but not as large as our encephalization quotient (ratio of brain size to body size).
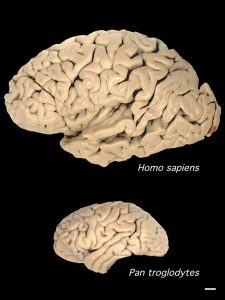
Brain size relative to body mass is one measure of difference but more important than this are organizational differences as measured by neuron count. The human brain is much smaller than that of an elephant or whale, but we have far more neurons in our cerebral cortex than any other animal. A Brazilian neuroscientist, Suzana Herculano-Houzel, developed a new way to count neurons. She concludes that primates evolved a way to pack far more neurons into the cerebral cortex than other mammals, with humans appearing to have the most cortical neurons of any species on Earth. Numerous neurons endow us with greater computational power and cognitive abilities. We average about 16 billion neurons in the cerebral cortex. This is the seat of our most sophisticated mental abilities, such as self-awareness, language, problem solving, and abstract thought.
Brain architecture, not just size, clearly differentiates human and nonhuman brains. The previous chapter briefly mentioned how differences in brain architecture between chimpanzees and bonobos goes far toward explaining why these two very closely related species have such distinct social worlds. The same extends to the comparison between humans and both chimpanzees and bonobos, but in an exponentially magnified way.
In many mammals, and indeed in many hominins, the frontal area of our brains is decreased in size. This is in large part because of post-orbital constriction, where just behind the eyes the cranial bones pinch inward, restricting the space available. In humans, however, we do not have post-orbital constriction, meaning we can have a large frontal lobe of our brain. Because the frontal area of our brains is large, we have prominent foreheads. Forehead shape is one distinctive trait that tracks the evolutionary change in brain development leading to fully modern humans and consequently this has been an important feature used by paleoanthropologists to track development in the human lineage, that and brain size, or how many cubic centimeters of cranial capacity a skull has. One role of the frontal area is for processing speech and language, and this is key for cultural transmission and social negotiation among individuals.
Teeth
Our teeth are markedly different from those of other apes. Like chimps and bonobos our dentition relates to our omnivorous diet which required generalized teeth. We can eat almost anything, but usually focus on the highest quality food, the stuff with the greatest payoff (we are optimal foragers!). Since we process this food prior to eating it and have being doing so for more than a million years, our teeth are not as robust. Fortunately, our teeth have thicker enamel than chimpanzees and bonobos, which is a benefit when you need to use them across many, many decades.
One of the most notable dental distinctions is that we have small canines compared to other apes, including even the somewhat more peaceful bonobos. They are puny by comparison and also incisor-like. With small canines there is no longer a need for a diastema; that trait disappears in human evolution. Primate canines play a social role involved mainly in male-male competition over access to females. The small canine size in human males is thought to be related to greatly reduced male-male competition for females during our evolution, dating back to more than 4 million years ago as evidenced by one ancient species of hominin (Ardipithecus ramidus) who had reduced canines (not as small as ours but far from chimp like). This trend continued such that when specimens in the paleontological record are recognized as belonging to the genus Homo, canines are greatly diminished.
Teeth are supported within the bone of the jaw and maxilla so if teeth change in morphology it is natural to expect some alteration of their support structure as well. One of the most notable of these was the change in jaw/maxilla shape from one that was like an elongated U in chimpanzees and bonobos (and other great apes) to a rounded shape that is more C-like in humans, like a parabolic arch. There was also a significant change in jaw size, which resulted in ever less projection of the face (less prognathism). With modern humans there is essentially no projection of the bones that house the teeth (upper teeth in the maxilla, lower teeth in the mandible).
Other Distinctive Traits
Human females are sexually receptive throughout the year rather than just during estrus and they have what is known as cryptic ovulation or hidden estrus. With some apes and monkeys, females will display swellings of the genital and perineal skin, a clear sign they are ready to mate. With such a visible sign, male-male competition always intensifies. Curiously, these sexual swellings do not seem to be consistent in the primate order, with some species possessing them and others not. For instance, chimpanzees and bonobos have estrous swellings, while gorillas, orangutans, and humans do not. While we cannot definitively say whether human ancestors did or did not have estrous swellings, the more or less continuous female sexuality and cryptic ovulation was likely a key part of human evolution that helped to dampen down male-male competition. This would have stabilized social interactions and allowed cooperating multi-male multi-female groups that perhaps consisted of bonded male-female families.
Another distinctive human trait among the greater apes in the high degree of male parental investment. Humans are more like many birds in this regard. Amongst chimps and bonobos, the male role beyond insemination is comparatively minimal when it comes to direct investment such as food provisioning or infant care. They do play a role in territorial defense that can be seen as indirect investment, though it is difficult to conclusively attribute this behavior to infant care. In modern humans, males can directly invest in many considerable ways, especially in nutrition to mothers and offspring, but also in childcare and instruction. The food acquisition part is exemplified by the common human pattern of a sexual division of labor in food-getting and food sharing. This goes hand-in-hand with adult males and females commonly forming pair bonds, the uniting of sexual and economic realms, to split the work load and commonly specialize in certain food-getting activities. Based on ethnographic and archaeological evidence of modern human societies, we can speculate that these behaviors were part of our evolutionary history that enhanced survival and reproductive success, though we cannot know for certain.
Humans have learned and culturally patterned behavioral flexibility, not only for the making and using of tools but also for cooperation and organization to achieve species survival. This makes us highly adaptive since we can respond quite rapidly to changes in the environment wrought by nature or ourselves. Humans have cumulative cultural evolution, which means that tools or techniques that work, i.e., that accomplish things successfully, get retained and often improved upon; they are not lost or forgotten about. Through this process human societies accumulate successful modifications and lucky errors over generations.

Cumulative cultural evolution builds bodies of knowledge that improve over centuries and millennia as this information is passed along from one generation to another and among different social groups.
- Knowledge about animal behavior and about edible plants.
- Knowledge about how to transform otherwise inedible or poisonous plants into food.
- Knowledge about how to capture fire and keep it going and eventually how to make fire from scratch.
- Knowledge about how to manipulate the physical world, initially with brute force and eventually with fire, to make entirely new substances like pottery, glass, and metal.
It is cumulative cultural evolution that has allowed us to be the only primate to hunt large animals, those greatly exceeding our body size; to survive at extremely cold temperatures and thus spread into environments unusable by apes or monkeys, including expanding to the western hemisphere where no apes existed; to survive ocean crossings and thus populate all remote islands of the world. Other animals have select habitats that they can live in, but culture has allowed humans to thrive in every corner of the world. Ultimately, who knows for certain since there are some extreme biological constraints, but it seems possible that cumulative cultural evolution will eventually enable humans to exist on other plants besides earth.
Even with smarts, any individual human would be hopeless stripped of our culture. If you were lost in the woods without matches, could you start a fire? If you were in a desert without water, could you find some? Would you be able to make a tool that could kill an animal for food or know where to forage and what was safe to eat?
The Downside of Large Brains
It is abundantly clear that in the course of human evolution there was strong selective pressure for increased brain size. It became a key trait responsible for why humans have been so successful. Thinking power really paid off. And any mutations that increased brain capacity and thinking power, such as neuron count would be selectively retained.
Can adaptations have downsides? Negative consequences? Yes! Adaptations are usually the result of some compromise. There are almost always evolutionary trade-offs. An adult human brain is roughly 6 times larger than expected for a mammal of our body size. This became a problem in our evolutionary history because selection for larger brains occurred after selection for bipedalism had narrowed the birth canal. Obligate two leg walking was present around 6 million years ago, well before the emergence of the human lineage—before the genus Homo.
The famous skeleton known as Lucy, an Australopithecus afarensis, was a habitual biped, meaning she was good at walking bipedally but also retained traits for life in the trees. Though Lucy had a slightly larger brain then chipanzees, it would still be considered small brain in comparison to ours. With bipedalism, there is a reshaping of the pelvis, constricting the birth canal. Though this may not have been as problematic for Lucy, as A. afarensis crania are not huge, in humans, giving birth is a hard and painful process because of needing to pass large heads through small birth canals. With selective pressure for larger brains in the human lineage, there developed an evolutionary compromise for infants with considerably underdeveloped or immature brains relative to adult brain size. The fetal brain at birth cannot be larger than about 400 cubic centimeters, otherwise the baby will not pass through the average female birth canal and death for child and mother can result. Human brain volume at birth is around 30% of adult size. This means that there is considerable brain growth during the first 2 years of life. This delayed maturation is a life history trait that may have been present even as far back as Lucy. Vital things for survival like walking and speech just have to wait while the brain grows & matures. This means that humans have the longest dependency period of any primate. Offspring require extensive parental care. The dependency period has only lengthened as our societies have become more technologically complex and specialized.
The Hominin Sequence
Humans are unique among primates in many respects, or at least we represent one far end of the continuum. Yet not all too long ago, there were other human-like species on the planet. Some of these species occupied the same or similar ecological niche and as such were competitors for space and resources. Just 40,000 years ago, Neanderthals still existed, but they went extinct soon thereafter. The small human species nicknamed the hobbit (Homo floresiensis) inhabited an Indonesia island until around 60–50,000 years ago. Two million years ago in Africa several different bipedal hominins lived. Some had no direct relationship to us but represented separate branches of human-like species. Despite all the diversity once present, we (Homo sapiens) now stand as the lone surviving representatives of what was a diverse hominin line. Many of us, though, carry a genetic trace of now extinct Homo species.
The term hominins includes all human ancestors back to our split from apes some 6.5-9.3 million years ago. Despite our close genetic relationship to chimpanzees and bonobos, there are many characteristics that set us apart from these other hominoids. These traits justify a separate designation for humans. It is quite obvious that the small fraction (<2%) of a difference between us and chimps/bonobos had a vast impact in evolutionary terms.
Geologic Time and Environmental Change
It’s important to have some understanding of the time period during which humans evolved. The key time range occurs during the Pliocene and much of the Pleistocene. This is the interval from just over 5 million years ago up to about 12,000 years ago. An informal short-hand designation for this interval is the Plio-Pleistocene. The Pliocene covers the first portion from roughly 5.3 to 2.5 million years ago. The Pleistocene covers the last portion and extends until just under 12,000 years ago. The latter part of the Pleistocene after about 200,000 years ago is less significant for the topic considered here since by then fully modern humans had evolved.
The Plio-Pleistocene was a time of pronounced environmental change. One critical factor related to hominins was geological and caused by the movement of tectonic plates. This caused the progressive rifting of East Africa, creating large lake basins, uplifted land masses, and high volcanic mountains. This dramatically altered the East Africa landscape from a “relatively flat, homogenous region covered with mixed tropical forest, to a varied and heterogeneous environment, with mountains over 4 km high and vegetation ranging from desert to cloud forest” (Maslin et al. 2014).
Adding to the pronounced environmental change was the alternating cycle of glacial and interglacial periods. During the ice ages, global temperatures dropped significantly. As such, water collected in massive ice sheets that advanced from polar regions and high elevations, covering up to one third of Earth’s surface and lowering sea levels by hundreds of feet. The warmer interglacial periods saw the retreat of ice the rising of sea levels and changes in flora and fauna across wide swaths. Throughout the Pleistocene there was ever increasing climatic fluctuation and environmental instability.
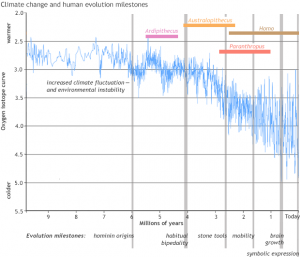
The above graph plots the oxygen isotope ratio for the past 10 million years. The ratio of oxygen-18 to oxygen-16 provides a proxy measure of global temperature. This proxy is obtained by measuring oxygen isotopes in the shells of microscopic ocean creatures called foraminifera. Ocean water has extra oxygen-18 during an ice-age, so the shells that form in the ocean at that time also have more oxygen-18. By measuring the amount of oxygen-18 in single shells from different intervals of time scientists can tell when glaciers were present and what the global climate was like. The oxygen isotope data indicate increasing variability in climate after 6 million years ago, becoming especially volatile during the last 2.5 million years.
Adaptive traits that typify humans are associated with the largest oscillations in global climate. This starts with the split in the ape line that separates species leading to us from those leading to chimpanzees and bonobos, which occurred before 6.5 million years ago. Habitual bipedalism then occurred in the hominin line, followed by cooperative groups, a decrease in tooth size, the making and using of stone tools, the scavenging of eating meat/marrow from large animals, wide ranging mobility (adaptive radiation to new continents), and ultimately brain growth and symbolic expression.
Some of these traits were originally thought to be hallmarks of the lineage that led directly to us, such as stone tools; but the evidence for this now dates back to roughly 3.3 million years ago, well before the appearance of the genus Homo and evidently associated with Australopithecines. And stone tools are simply those that survive in the archaeological record and were likely preceded by tools made of organic materials such as wood. The onset of long-endurance mobility and obligate bipedality is associated with Homo erectus, the first hominin to leave Africa and the first to have limb proportions approximating ours. Rapid brain enlargement is something that began about 800,000 years ago; over the course of human evolution, brain size tripled and the majority of this expansion occurred after 800,000. Symbolic expression, innovation, and cultural diversity, started to occur after about 100,000 years ago and has accelerated ever since, as cultural innovations built one upon the other.
Rick Potts thinks that it was the highly variable environment that was a driving force behind the evolution of our mental abilities, which is what facilitated culture. Natural selection favored the learning of behavior because culture allowed for behavioral plasticity. Hominins most adaptable to change because of depending on learned behaviors were better able to survive and reproduce and ultimately this led to hominins that depended on culture.
Early Hominins
Divergence from the common hominid ancestor for humans and chimpanzees/bonobos occurred during the late Miocene before the onset of the early Pliocene. The timing on this is estimated by the molecular clock (or genetic clock), which is based on the ongoing evolutionary processes of mutation and recombination. These two biological processes generate heritable variation in genes, changes to the base pairs of DNA’s genetic code. Mutations accumulate slowly, so the mutation clock works best for ancient events, like evolutionary splits between species. The recombination clock works better for events within the last 100,000 years. Based on the number of mutation events that exist between humans and chimpanzees, the timing for when our last common ancestor existed is between 6.5-9.3 million years ago.
Fossils of early hominins dating to after the divergence from chimpanzees/bonobos are extremely rare and quite fragmentary. This is the general trend in paleoanthropology: the further back in time one looks, the more difficult it becomes to find evidence. Bone fossilization itself is a rare event for terrestrial lifeforms since it requires burial in a proper depositional setting. Then there is the issue of long-term preservation of that rare fossil evidence: erosion and other geological processes constantly destroy evidence, winnowing down what was a small sample to begin with. Then comes the rare chance for discovery, something that is almost totally fortuitous, even for professional seekers who know how to find fossils and where best to look.
Since fossils from ancestral hominins are rare, fragmentary, and often in poor shape, our understanding of human evolution is constantly changing and improving, with each new find. New finds can overthrow past hypotheses or models about relationships between different species. There is also constant debate about how different finds should be classified: is this find part of OUR direct lineage or a tangential lineage? These debates can sometime be acrimonious. There was and continues to be a tendency to designate a new species for any somewhat anomalous new find. Finding and naming a new species is more noteworthy for a scientist, a quicker path to fame than just documenting yet another case of a preexisting species. New finds often garner more research funding and larger grants of research money.
The earliest hominins so far identified are classified as members of three distinct genera; from oldest to youngest these are Sahelanthropus, Orrorin, and Ardipithecus. Much remains to be learned about the morphology and environments of these taxa, so there are no definitive conclusions at this time about such issues as taxonomy and the environments of adaptation that selected for human-like traits, such as bipedalism and small dentition, including reduced canines.
Sahelanthropus tchadensis
The earliest known hominin is called Sahelanthropus tchadensis, discovered in Western Chad. The suggested age is around 7 million years ago but unfortunately the region lacks the volcanic ash layers that provide such excellent radiometric dating in places like the rift valley system of east Africa. While this species is known primairly from cranial elements, recent fossil finds of femora (plural of femur) have been assigned to this species. An almost complete skull combines a mix of human-like and ape-like traits. Hominoid (ape traits) include large brow ridges, small brain, wide face, large sagittal crest, U shaped dental arcade, marked postorbital constriction. Hominin (human) traits include smaller canines with thick enamel, more vertical face, no diastema, and more central positioning of the foramen magnum. The latter indicates a more bipedal stance and mode of locomotion that was perhaps habitually bipedal (not a knuckle-walker like chimps/bonobos). The femur is not much more conclusive as it lack many of the features (specifically the proximal epiphysis, or hip joint) that would be used characterize bipedal locomotion. What can be ascertained is that the size of the femur is consistent with other fossil hominins, but the morphology is most correlated with the genus Pan (chimpanzees and bonobos). Thus, the femur is definitely not like that of a habitual biped, meaning bipedal leg morphology had not yet evolved with Sahelanthropus tchadensis.
Orrorin tugenensis
Appearing soon after Sahelanthropus, living around 6 million years ago, is Orrorin tugenensis. Orrorin was perhaps a descendant of Sahelanthropus, but hard to know for certain right now because an Orrorin skull has yet to be found. Currently, there are just jaw fragments and isolated teeth, as well as a few fragments of arm and leg bones. This species also had a combination of ape-like and human-like features. A femur head is an important find since it is derived in its morphology (different from any fossil apes before it) and indicates advancements in bipedal capabilities. Orrorin’s teeth are also small with thick enamel, similar to those of modern humans.
Ardipithecus
The next member of the early hominin finds includes the genus Ardipithecus, with two species recognized: A. kaddaba and A. ramidus. Both species come from late Miocene layers in Ethiopia. The earlier species, A. kaddaba, is known by relatively few fragmentary remains whereas A. ramidus is known from a modest number of specimens including a partial skeleton nicknamed “Ardi.” The mixture of ape-like and human-like features is quite clear, because of the partial skeleton. Human-like features include small, blunt canines, lack of the ape dental arrangement that maintains tooth sharpness for edges of the upper canine and lower first premolar (the C/P3 honing complex), a more centrally positioned foramen magnum along with other characters indicative of habitual bipedalism. Ape-like features include a fully opposable big toe, an ape-like lower pelvis, relatively equal fore- to hind-limb lengths, and long, curved fingers and arm bones that indicate it retained aspects of arboreality.
Middle Hominins: The Australopithecines
Australopithecines includes all species in the genus named Australopithecus as well as those more robust forms included in the genus named Paranthropus. The Paranthropus species are also known as the robust Australopithecines because they had thicker bones, significantly more massive jaws and teeth, and a sagittal crest on their skulls. The species of the gracile group are in the genus Australopithecus and lack sagittal crests and have smaller jaws and teeth than the Paranthropus species, indicating quite distinct diets between these two groups. Paranthropus likely subsisted mainly on tough fibrous plant foods similar to the diet of modern gorillas, whereas the Australopithecus species likely had a higher quality diet based on softer fruits and animals.
The first australopithecine find occurred in 1924. Since then, there have been hundreds more finds. All species of both the robust and gracile groups were capable of bipedalism but with brains only slightly larger than those of apes. In evolutionary terms this means that functional bipedalism occurred well before there was natural selection for greatly expanded cranial capacity in the human line.
There is unquestionable anatomical evidence for bipedalism and the skeletal evidence is rather proof positive. But if doubts remained, Mary Leakey put them to rest with the discovery of fossilized footprints at Laetoli in Tanzania. The volcanic ash that the footprints were made in is directly dated to 3.6 million years ago. There is absolutely no doubt about bipedal locomotion. It is presumed that Australopithecines left these prints. The prints clearly show individuals walked on two legs quite similar in manner to us (see the video below). Moreover, they seem to have traveled in a small family group.
Despite being fully bipedal, Australopithecines were also proficient at clambering around in trees. Evidence from arm and hand anatomy strongly indicates a continuing arboreal adaptation, at least to some extent. They seemed to have engaged in plenty of two-legged walking on the ground mixed with tree life, likely for both food and for safety. Being rather small in stature (both Australopithecus and Paranthropus species), Australopithecines would have been prey to many of the large predators of the African savanna and woodlands. The giant hyena known as Pachycrocuta that existed alongside hominins was close to lion size, with a mouth capable of fully enveloping a hominin’s head and removing it. Like today’s hyenas it was a pack animal, so all the more fearsome. Early stone tools would have been no defense against such an animal.
The robust Paranthropus species were not in any direct line leading to the genus Homo and eventually to us (the Paranthropus name means ‘parallel to’ or ‘alongside human’). It was from one of several gracile Australopithecus species that our genus likely evolved, although which species remains a hotly debated topic. There are at least five well recognized Australopithecus species and even more depending on what specialist you talk to. All have smaller dentition and lighter facial and dental muscles than the robust Australopithecines.
There are at least three successive Australopithecus species recognized for east Africa, with fossil finds occurring from Ethiopia south to Tanzania: A. anamensis, followed by A. afarensis, and then A. garhi. In South Africa there are at least two successive species: A. africanus and A. sediba. The east African specimens have much better chronological control because of the occurrence of volcanic ash layers that can be dated radiometrically. Finds in South Africa occur in a different depositional environment, that of limestone caves. In many cases the bones got deposited there as the food scraps of predators such as leopards, or in the case of the Taung child an eagle. Each of these five species are briefly characterized below.
East African Australopithecus Species
Australopithecus anamensis
Australopithecus anamensis is the earliest species of this genus. It lived some 4.2 million years ago. It might be descended from Ardipithecus ramidus and was perhaps ancestral to A. afarensis, although there is also evidence of temporal and spatial overlap between these two species. As with the earlier Ardipithecus ramidus, A. anamensis has traits intermediate between apes and humans.
Australopithecus afarensis
A. afarensis has the largest sample of fossil remains of any Australopithecine species, with the remains of more than two dozen individuals from Laetoli, Tanzania and at least 35 individuals from Hadar, Ethiopia. There are also the fossilized bipedal footprints in volcanic ash of several individuals, with the ash well dated to between 3.8-3.6 million years ago.
One of the most complete hominins from this time period is the A. afarensis skeleton known as “Lucy” that dates to about 3.2 million years ago. The name came from the Beatles song after an all-night party following the find. This skeleton is about 40% complete and includes many critical features such as a full femur, knee and ankle joints, arm and wrist joints, and skull portions including most of the lower jaw. This skeleton has a human-like pelvis, femur, knee joints, feet, and head placement with a central foremen magnum. But these are coupled with long apelike arms, long grasping fingers, and flexible feet for walking or climbing (the above video shows Lucy’s traits). This combination of features appears characteristic of Australopithecus for nearly 2 million years. It afforded an ability to move in diverse habitats by changing the degree of reliance on terrestrial walking and arboreal climbing. Such flexibility also seems true for earlier and perhaps ancestral hominins such as Ardipithecus ramidus.
Australopithecus garhi
This individual continued the evolutionary trends of the Australopithecines in eastern Africa. Compared to A. afarensis, A. garhi had slightly less prognathism (the jutting out of the lower half of the face), but had slightly larger dentition than A. afarensis. Its femur showed signs of a habitual biped, but its arm length was still greater than its leg length, suggesting arboreal tendencies.
South African Australopithecus Species
Australopithecus africanus
One of the most famous of the A. africanus finds is known as the Taung child. Discovered in 1924, Taung child was actually the first Australopithecus identified and the first hominin ancestor found in Africa. Dating to between 3.6-2 million years old, A. africanus has a small, ape-like brain case but slightly smaller, more human-like teeth. It was these features that led scientists to eventually accept that a large brain was not the defining characteristic of our earliest ancestors. Like the other Australopithecines, A. africanus had more ape-like limb proportions, meaning they still had a heavy reliance on arboreality. But, the inner structure of the femur bones, which resembles that of humans, indicates that bipedalism was definitely part of their repertoire as it was able to bare loading weights of an upright posture.
Australopithecus sediba
Australopithecus sediba is another recent addition to the Australopithecus lineage. The first specimen was found by Lee Berger’s nine-year-old son in 2008. The boy was visiting his dad’s dig site in South Africa and he literally stumbled on a bone which turned out to be a clavicle embedded in rock. When his dad turned the rock over, there was a mandible with a canine. Because of various skeletal features in both the cranium and lower bones, Berger suggests that A. sediba may be a transitional species between the A. africanus (the Taung Child) and later Homo habilis or even Homo erectus. This hypothesis is disputed by many specialists and the exact link between the genus Australopithecus and Homo has yet to worked out. A. sediba shows very clear bipedal morphology in its legs and hips, possible more derived than any of the other Australopithecines discovered thus far. But, it still clearly has adaptations for arboreality, meaning it still is not an obligate biped and forested areas are still important to its survival.
Genus Homo
Human evolution was once envisioned much like a simple twig, progressing in relatively straightforward fashion up the stem to us, the flowering bud at the apical end. We now know that it was a massively bushy tree, with major branches that had nothing to do with us. One of these major branches is our genus of Homo but even that branch is messy, with dead ends or extinctions. Moreover, the tree analogy is probably inappropriate, since human evolution was more like a heavily braided stream, with separations and then mergers, and then further separations. Some of the “dead ends” such as Neanderthals, contributed to the overall end result. This is why many modern humans have some Neanderthal DNA. If you get your DNA tested you can learn how much Neanderthal contribution occurs in your genome; it might be 0% or up to 4%.
After about 2.5 million years ago is when there are hominin fossils that get classified as part of the human genus Homo. The first of these to be recognized is named Homo habilis or the “handy man.” Subsequently Homo rudolfensis was recognized, although some think that this species merely represents a range of normal variability (just as any species has some variation) and that it is actually just H. habilis. Exactly which previous species gave rise to the Homo genus is still in debate. Some say A. africanus, some A. sediba, some say other species. This debate will eventually be resolved with more field work; it cannot be resolved by arguing from the fossil specimens currently in collections.
Finds of new Homo species are still being made and there are doubtless more to come. Homo floresiensis, the “hobbit,” was discovered in 2003, H. naledi was discovered in 2013, and H. luzonensis was discovered in 2019. All of these species date relatively recently in our evolutionary history: H. floresiensis from 100,000 to only about 50,000 years ago, H. luzonensis from 67,000 to 50,000 years ago, and H. naledi from about 335,000 to 235,000 years ago. All of these new species are small-brained, more like Australopithecines, yet they had many human-like anatomical aspects of the hand, foot, lower limb, dentition, and cranium. It is strikingly clear that there was no simple linear progression towards modern humans, and different morphological forms overlapped in time and possibly even competed with each other. Instead of a straight or linear progression, evolution appears to be more mosaic in nature, experimenting with different pieces at different times to see what works in the moment.
Early Homo
Homo habilis and Homo rudolfensis
The three earliest members of the genus Homo include H. habilis and H. rudolfensis at the base, beginning sometime about 2.4 million years ago, with H. erectus starting about 2 million years ago. Some argue that H. habilis and H. rudolfensis simply represent the range of variability that can be expected for a single species. The cranial capacity of both H. habilis and H. rudolfensis was about half of what you have. H. rudolfensis is thought to have a cranial capacity just slightly larger than that of H. habilis on average. However, the specimens identified as H. rudolfensis may have also had a larger body (there are no definitely identified postcranial remains); thus the size difference could be easily interpreted as sexual dimorphism – much like average size difference between males and females in modern humans. Homo habilis and H. rudolfensis both had reduced molars and premolars compared to Australopithecus.
Based on the cranium, H. habilis looks more human than ape when compared to Australopithecines. It has a larger brain along with more human-like dentition than Australopithecines. Directly tied to smaller teeth and smaller chewing muscles was less jaw projection or prognathism; H. habilis had a flatter faced than Australopithecines. But below the head, H. habilis was less distinct from Australopithecines. Homo habilis had disproportionately long arms compared to us (the subsequent H. erectus was proportioned more like us). Homo habilis had powerful hands to go with those long arms. Powerful hands and the long arms suggest the continued importance of being able to escape the ground by climbing trees, which had to have been much safer place after the sun had set. Strong hands were also evidently suited to making and using stone tools (Oldowan technology) and likely other tools as well, such as those made of wood.
Homo erectus
Homo erectus occurred later than the previous species, beginning sometime around 2 million years ago, and overlapped with them for perhaps a half million years despite also being their likely descendant. H. erectus represents the first expansion of hominins out of Africa leading to the establishment of human-like species in Asia and Europe. Because the species name H. erectus was first used for finds in Asia (Java and China), some specialists restrict that label to Asian finds and refer to similar looking finds from Africa as Homo ergaster.
The skeleton known as Turkana Boy is an example of H. erectus (or H. ergaster if you prefer) who lived in Africa about 1.5 million years ago. This is the most complete H. erectus skeleton yet found. He was tall and slender with limb proportions very similar to modern humans. In adulthood he would have stood at least 6 feet tall, but he died before this. He was much taller than H. habilis and Australopithecines (both about 4 feet tall) and since he died before reaching adult hood, the height difference would have been even more pronounced.
Even though the H. erectus body anatomy was very similar to ours, cranially there are some obvious differences. Though their brain was larger than before, the H. erectus brain was smaller than that of modern humans and evidently organized differently because it lacked the greatly expanded frontal lobe seen in modern humans. Their skull was long, low, and thickly walled, with slight post-orbital constriction. H. erectus skull also had pronounced brow ridges and a low ridge running along the crest of the skull; another ridge of bone ran horizontally along the back of the skull. The trend for a flatter face first seen in H. habilis also continues with H. erectus. This follows the prominent evolutionary trends among the genus Homo for a significant reduction in the size of teeth and jaws, a reduction in enamel thickness – H. erectus had the thinnest enamel of any hominin we have discussed so far – and muscles that work them and hence a flattening and reduction in facial features related to chewing. These traits are all linked together and reflect a change in diet to softer food items and to the probable processing of hard foods with tools and/or fire to make them softer.
Human Evolution and Tool Use
Stone tools and other cultural adaptations are key in our evolution. Stone tools provide a record of changes in our mental and physical abilities over the past 2+ million years. Homo habilis was once thought to be first hominin tool user and tool use was thought to be a unique part of the genus Homo. Tool use made us distinctive and gave us the competitive edge over other hominins and other animals. Such a notion occurred before Jane Goodall documented tools use among chimpanzees and well before the documented use of stone tools by some New and Old World monkeys.
We now know that some species of Australopithecus also used stone tools. Stone tools occur in deposits dated well before the emergence of the first identifiable Homo species. The earliest stone tools date to about 3.3 million years ago in Ethiopia and are likely associated with Australopithecus afarensis.
It is becoming clear that using stone tools was not the most important thing in our evolutionary history, despite some claims to the contrary. Other behaviors could have mattered far more, such as greater cooperation within the group and between groups. This could have had a huge advantage for our species as we had to compete with other organisms for finding food and occupying ecological niches.
Perhaps most significantly, was the use of fire. Using fire for cooking made foods not only softer but more digestible and with more nutrition made available. The use of fire “captured” from the wild and ultimately controlled (the ability to start one when need arose) was a major step forward for humans. This is still something that sets us apart from all other animals and H erectus pioneered this unique behavior. Aside from the nutritional benefits of cooking food, fire help to protect from predators and was likely key to expanding into cold environments because of the warmth it provided.
The Oldowan Tradition
The first recognizable earliest stone tools associated with the genus Homo, thus representing the most diagnostic and earliest of human culture, is the Oldowan Tradition, named after Olduvai Gorge in Tanzania. Oldowan technology consists of pebbles about the size of a baseball or softball with flakes removed from them on one end and the simple flakes detached from such cobbles. They are associated with H. habilis and H. rudolfensis. A simple technology like this persisted for a very long time. Even after the evolution of fully modern humans capable of making sophisticated stone tools, humans might have also created simple Oldowan-looking tools for some expedient task.
The Olduvai Gorge is a massive ravine in the Great Rift Valley of eastern Africa and is one of the most important field areas for hominid evolution. As far back as 1913, geologists began to study and describe the various geological beds in numerical order. The geological layers are rather straight forward with relatively horizontal strata, like a layer cake, that formed in and along brackish lakes. Moreover, the layers are interspersed with beds of volcanic ash (tuffs) that provide for accurate radiometric dates, with the oldest at 1.9 million years ago. The Oldowan stone artifacts in the gorge seem to originate from substantial distances away from the lake, suggesting advance planning by the Homo species. They thought about what tools that would be needed for some tasks, went and collected them, then brought them to the place of use where the tools were prepared, used, and discarded.
What is a geological bed?
- A deposit of sediment of variable thickness that is distinct from underlying and overlying depositional layers; they are often called strata.
- At Olduvai Gorge these beds are representative of lake and lake margin deposits and many beds are separated by volcanic ash layers (tuffs) that can be dated.
- If stone artifacts or human remains occur within a tuff layer, we can say that such finds have the same age as the tuff.
- When stone artifacts or human remains are found below or above a dated tuff layer, we can say that such finds are older than or younger than the tuff.
Throughout her research in Olduvai Gorge, Mary Leakey defined the Oldowan Tradition (or culture) as primarily consisting of pebble “choppers.” Today these are defined as “cores” and they comprise less than 30% of a total assemblage at the various sites; flakes detached from those cores comprise 70% or more. Mary Leakey considered the cores to be the true tools and the flakes, or debitage, to be manufacturing debris. Flakes were detached by striking cores at the proper angle using a hard hammerstone for direct percussion. These days archaeologists recognize that the flakes are in many cases also tools, used in cutting through animal hide and likely other materials, slicing off meat, and scraping. Many of the stone cores likely also functioned as tools, like Mary Leakey thought, such as for cracking open marrow-rich long bones.
The Acheulean Tradition
H. erectus had a more sophisticated flaked stone tool technology than H. habilis, one that archaeologists call Acheulian technology. Acheulean technology consists of more refined stone tools than those made and used by H. habilis. Acheulean technology emphasized what archaeologists refer to as bifaces, which simply means cores that are flaked on two faces to make them somewhat flat and thin. The Acheulean bifaces, often referred to as handaxes, are somewhat teardrop-shaped with a sharp point and sharp adjacent lateral edges. The rounded “back” of the tool might be less flaked or have blunt edges and is assumed to be where H. erectus held them when in use. These bifaces are associated with H. erectus and also later Homo species and the bifaces become more refined thru time, especially after about 1 million years ago. The Acheulean name for this flaked stone technology is derived from a location in France because tools of this form were first recognized there before being discovered in Africa, where the technology originated. As with Oldowan tradition, archaeologists have traditionally emphasized the core tools, those handaxes, as the beginning of the focus on technology. But, just as with Oldowan the flakes that got detached while making the bifaces were likely important tools as well, at least the larger flakes.
This new more refined tool form is unlikely to be the key reason why H. erectus expanded across much of the rest of the Old World. Rather, the tool form is a reflection of increased mental capacity and greater ability to plan and execute tasks and certainly those aspects were key. Through time the species of Homo increasingly used more and more diverse types of tools. The ones we see are mainly those that preserve, which means those of stone, but they probably reflect just the tip of the iceberg in technology. The earliest wooden tools found are spears from Schöningen Germany dating to more than 300,000 years ago. These likely reflect an exceedingly long tradition of tools made from perishable artifacts, which is what chimpanzee technology consists of. Tools that preserve well, those of stone and bone/ivory, become increasingly complex thru time. Brain size also increased but it is not necessarily tool use that is driving this process. The smarts that can lead to better tools are also engaged in most other aspects of human behavior that lead to better odds of survival and reproductive success.
Hunting or Scavenging?
Some of the initial speculation about why H. habilis made and used stone tools was because it allowed them to kill animals. Stone tools got linked to the killer ape hypothesis, which envisioned early humans not as scavengers but as active hunters. This conviction was on display in the remarkable opening scene of Stanley Kubrick’s masterpiece film 2001: A Space Odyssey. Robert Ardrey popularized this notion in his book African Genesis: “Man had emerged from the anthropoid background for one reason only: because he was a killer.”
This highly speculative account is now discredited! While hunting eventually became central to human survival and our ability to spread around the world, this was not the case early in our history. Stone tools for hunting is a case of later day function applied to the origin of tools. Stone tools were eventually critical for the hunting of large game animals, but initially Oldowan technology was about scavenging meat and marrow from dead carcasses. The benefit of even simple stone tools, which is what Oldowan flakes and pebble choppers are, becomes quite obvious when you think about what you would do if you happened upon a dead rhino and were exceptionally hungry.
Hominins lack the teeth or claws necessary to rip into thick animal hide or crush massive long bones. Vast food resources on the savannas of Africa were inaccessible to hominins without stone tools. If a H. habilis group happened upon a dead rhino they could have done very little until lions or hyenas had ripped into the carcass. The ability to create sharp stone chips and sharpened chunks of stone meant that hominins could get to both meat and marrow. Tools meant better access to food resources, so greater survival and reproductive success. And those same scavengers could easily take down an early hominin, so speed at removing food from a carcass was also essential to avoiding predators.
Hunting large game with an Oldowan pebble chopper seems farfetched, but how might we know who was killing the large game found in association with H. habilis remains or their tools? Priority of access is one key indicator that H. habilis was chiefly a scavenger; simply put, they did not get first access to meat on game animals. At many early sites, stone tool cut or abrasion marks made by H. habilis are on top of tooth marks made by carnivores. This means that carnivores got there first and indeed, they probably made the kill. In short, various lines of evidence on skeletal elements indicate that H. habilis mainly got whatever was leftover.
Briana Pobiner spent seven months investigating whether scavenging was worthwhile; did it have a sufficient payoff? She simulated passive scavenging in Africa by waiting until carnivores had eaten their fill and moved off. She documented how much meat and marrow was left on carcasses. Turns out it’s quite a lot! In her sample of lion kills of larger animals, mainly zebras, 95% of the bones had at least some flesh remaining, and over 50% had significant meat left. The average zebra hind limb contains almost 23 kilograms of meat, so when 90% is consumed, it could still yield over 2 kilograms of meat. That’s just 1 hind limb. An entire zebra carcass can yield almost 15 kilograms of scavenged meat. That’s about 60,000 calories or the rough equivalent of 107 Big Macs. Its enough for the entire daily caloric requirements of about 30 male H. habilis. And that excludes the calories available from crushing long bones to extract the fat-rich marrow.
Homo habilis was perhaps chiefly a scavenger, at least of large game animals. They may also have active hunted small game, especially those that were less mobile and easily captured. It’s also good to remember than H. habilis was also the potential prey of numerous dangerous predators in Africa. Early stone tools were little if any defense against the common large lions, saber-tooth cats, hyenas, and leopards that roamed throughout Africa.
Later Homo
The expansion of H. erects out of Africa was part of an adaptive radiation for Homo, one that led to the diversification of species. This is a common occurrence in evolution, as species spread widely and then become reproductively isolated to varying degrees and from various influences (genetic drift). Various Homo species are recognized in the Old World as descendants from H. erectus, some of which are well known from skeletal remains (e.g., Neanderthals) and some mainly from genetic evidence from rather than their rare bones (e.g., Denisovans). Others, such as Homo heidelbergensis are known from some skeletal material, mainly skulls, but not DNA.
There is much disagreement and debate about how all the various pieces of this rather late human evolutionary puzzle fit together, likely in part because so many of the puzzle pieces are still missing. The muddle in the middle is a commonly used phrase for this interval in human evolution. The fortunate aspect is that with some of the more recent remains, DNA extraction is possible, which allows small skeletal fragments such as a single digit to provide significant information. This would not be the case based just on morphology.
Neanderthals
If you ever worried about an image problem for yourself, then you just haven’t lived as a Neanderthal. They have such issues in spades. Brutal killers is one portrayal, as in the Frank Frazetta painting shown here from 1966. That is a nightmare that will not just wake you but keep you from going back to sleep! He was reflecting some of the speculations about this cousin of ours that were current at the time. This image has not totally gone away as exemplified by the pseudoscientific account of Danny Vendramini and his Neanderthal predation speculation.

A far different image is portrayed in the 1981 film Quest for Fire. Rather than brutal killers, that film depicts Neanderthals as not all that smart. They are on a quest for fire because their fire went out and they don’t know how to start one. Neanderthals are shown as culturally inferior, with a simple technology, relative to that of the modern humans that they eventually encounter. It is only in the last decade or two that research has started to erode this idea of Neanderthals as culturally inferior. After all, they survived in Europe and parts of western Asia for a few hundred thousand years.
The first Neanderthal skull and bones were discovered in a cave in Neander Valley of Germany in 1856. These and other Neanderthal discoveries in Europe led to the idea that these ancient Europeans played an important role in modern human origins. It is now clear that while Neanderthals were surviving in Europe and western Asia, fully modern humans were evolving in Africa. Fully modern humans ultimately expanded out of Africa in yet another wave of migration and ultimately assimilated and replaced Neanderthals and other human species, such as the Denisovans in Asia.
It is possible that Neanderthals, Denisovans, and likely other Homo species, such as H. heidelbergensis, were descendants of that first adaptive radiation of Homo erectus, although the exact details of this are still being worked out with both fossil finds and DNA research.
Key evidence for how Neanderthals relate to modern humans comes from mitochondrial DNA (mtDNA), a special type of DNA inherited only from the mother. mtDNA was first extracted in 1997 from the Neanderthal type specimen, the remains that had been discovered in 1856 and that sat ready for study in a museum display (an excellent demonstration of the value to saving collected materials in museums). The extracted mitochondrial DNA was quite different from modern humans. A comparison of Neanderthal DNA to that of modern humans and chimpanzees found that most modern human sequences differ from each other by about 8 substitutions on average. Neanderthal and modern human sequences differ by 27 substitutions. Modern human and chimpanzee sequences differ by about 55 substitutions. Based on this, the last common ancestor of Neanderthals and modern humans dates to around 600,000 years ago. This is the approximate time of divergence between Neanderthals and the lineage that evolved into us. Since this initial study by Svante Paabo, many more samples of Neanderthal mtDNA have been recovered. The newer studies have replicated the original findings.
While the mtDNA provides one story about Neanderthal and modern human relatedness, nuclear DNA provides another. Svante Paabo and others have studied Neanderthal nuclear DNA. Neanderthals and modern humans have identical versions of several genes. The FOXP2 gene is one and this seems to be linked to the ability for language. Nuclear DNA establishes that Neanderthals and humans interbred on a few different occasions. Neanderthals had fair skin to aid with Vitamin D production and some had red hair (linked to reduced melanin production). Neanderthals contributed both useful (genes for immune response) and deleterious (blood clotting and depression) traits to modern humans. Research by Svante Paabo indicated that vulnerability to COVID-19 and the possibility of worse outcomes is linked to a DNA region evidently inherited from Neanderthals.
Neanderthals have gotten something of a makeover in recent years. Although originally stereotyped as dim-witted brutes with rudimentary culture, Neanderthals are now seen as thoughtful, artistic and spiritual—almost like us.
Neanderthals probably were capable of starting fire, but that and their other survival abilities did not matter in the long run. Neanderthals went extinct within a very short interval after the arrival of fully modern humans. Facts of the case are this: Neanderthals dominated Europe and the western part of Asia for at least 300,000 years. That is a successful adaptation by any measure. They successfully survived through intervals of glacial advance and retreat and much environmental change until they had direct competition for the niche that all species of the genus Homo occupy. Modern humans arrived in Europe roughly 45 thousand years ago. Neanderthals were gone only about ~5 thousand years later. They may not have been in direct conflict, though that could have occurred, but modern humans were better at getting food and probably had higher birth rates. Thus, Neanderthals were assimilated and out-competed in a niche in which they had been so successful.
Neanderthals provide a good example of a point we tried to make in the chapter on biological evolution: there is no such thing as a perfect adaptation. It all depends on the environmental context. Neanderthals were doing great, had been for some 300,000 years. What happened? The environment changed from a glacial to and interglacial period and a new competitor arrived: anatomically modern humans. With the environmental change, food sources inevitably changed. At the same time, humans occupied the exact same ecological niche as Neanderthals.
Archaeological evidence of cultural tradition, especially stone tools, also supports the temporary co-existence of two species in Europe and the Near East. Different tool making traditions also track the wave of population replacement as modern humans spread into the middle east and then Europe. Sites in Israel for example contain evidence of occupations by modern humans and Neanderthals overlapping for about 30,000 years. The separate cultures of these separate species are as follows:
- Mousterian (Middle Paleolithic) = Neanderthal
- Aurignacian (Upper Paleolithic) = Homo sapiens
The Mousterian Tradition
The Mousterian flaked stone tool technology is one that clearly emphasizes flakes rather than the cores that the flakes came from. The key diagnostic aspect of this technology, is that the core was well formed by detaching a whole series of preparatory flakes. The resulting shape of the prepared core would then allow for a desirable high utility flake to be detached. After removal of this large flake, the core was reshaped by flaking so that another flake of desirable shape and size could be removed. This process would continue until the core was too small to effectively remove a desirable flake. The number of iterations depended on the size of the original chunk of raw material, with large pieces allowing for the removal of more such desirable flakes.
This method of making flakes from prepared cores is also known as Levallois (lev-all-WA) technique. It was widely practiced by Neanderthals and took considerable planning, hand-eye coordination, and flintknapping skill to execute it well. The core shaping is all about producing optimal flakes of a given size and form. The preparatory flake detachments to shape the core and then to create the proper surface (platform) for striking the core, were essential to creating desired flakes.
Flintknapping Videos
If you want to learn more about how flaked stone tools are made, please watch the videos linked below. They are all in French without English subtitles but are the best ones out there for illustrating the making of an Acheulean biface, a Levallois core and flake (Mousterian Tradition), and the the subsequent Solutrean Tradition of tools made on blades.
Acheulean Biface
The video linked below shows the production of the Acheulean biface that is found in Europe and was chiefly made between about 400,000 and 100,000 years ago. It is a more refined version of the original Acheulean bifaces first made in Africa, which were all produced by hard hammer percussion alone: stone hammerstones only, no use of antlers as shown in this video. The innovation of using softer organic tools for percussion was a development that occurred after 1 million years ago. Soft hammer percussion allowed for thinner flakes to be detached, which created thinner and more symmetrical bifacial tools.
 Levallois Core & Flake
Levallois Core & Flake
The video linked below shows the careful preparation flakes that get removed for the creation of a proper Levallois core of the Mousterian Tradition. The preparation flakes are detached to create a core that has a humped up back side and are more gently convex top side. It is from this top side the the desired flake will be removed, one that has an elongated cutting edge. Note the careful preparation of the platform or striking surface, which has to have the appropriate angle and size for striking off the large desired flake. The planning and skill in the execution of this flaked stone tool technology should be quite evident. Neanderthals routinely did this feat of flintknapping.
 :
:
Solutrean Point Made on a Blade
The video linked below shows the fabrication of a Solutrean projectile point of the type made in portions of western Europe (modern day France, Spain and Portugal). On display are a few new flintknapping techniques innovated by modern humans. One of these is the production of long and narrow flakes, called blades. These are detached from highly prepared cores of specialized form. This was an elaboration of the Levallois technique that built upon the realization that by careful shaping of the core, the flakes detached will have desired properties. It should be obvious that the blade production is far more conservative of raw material than the Levallois technique. A single chunk of raw material can be used to detach dozens of straight sharp-edged blades compared with just a few ideal flakes with the Levallois technique. In this video the blades are struck off by direct percussion using an antler billet, but another innovation that occurred at this time was indirect percussion where a punch is held on the platform surface and then struck with the billet. This allows for greater control. Another flintknapping innovation shown in this video is pressure flaking, where a pointed piece of antler is used to “push off” small flakes. This allows for even greater control in the shaping of flaked stone tools.

Modern Homo sapiens
Fully modern humans, Homo sapiens, evolved in Africa after around 200,000 years ago. This is evidenced by fossils from sites in Ethiopia and in Morocco. One key find is from Herto, Ethiopia, dated around 160,000 years; these fossils look modern. The dating on when modern humans left Africa is still being worked out. It has generally been thought to have occurred around 100,000 years ago. More recent finds, such as those from a cave in Israel, are pushing this date back. This is always the case where the basic evidence, which consists of fossils, is always being discovered.
Holocene Migrations
Final Expansion – Oceania
- Melanesia: The islands around and including New Guinea; primarily named after the skin color of the people inhabiting these areas;
- Micronesia: Small, very old coral atoll islands;
- Polynesia: The three corners defined by New Zealand, Hawai’i, and Easter Island.
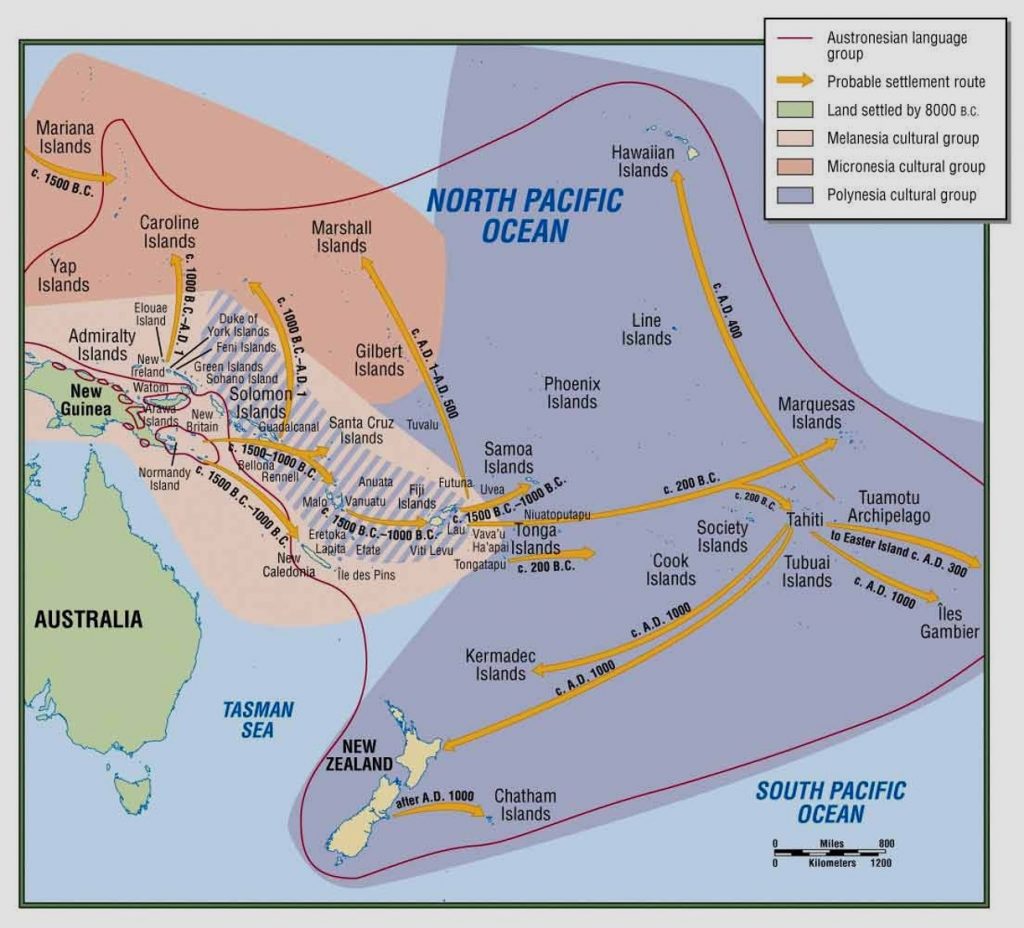
This is the final expansion of humanity over the planet was to spread into the islands of the Pacific region. After this expansion, all areas of the globe were occupied by humans, except for Antarctica. This expansion began from Asia and has been traced by examining the dates of a specific type of ceramic called Lapita. Lapita pottery has a time transgressive sequence that reveals the expansion of peoples into the Pacific. What that means is that by looking at the ceramics and the associated radiocarbon dates, it shows the expansion roughly from the Asian mainland to Polynesia with the earliest dates in the Asian mainland and the latest dates in the three corners of Polynesia – New Zealand, Easter Island, and Hawai’i.
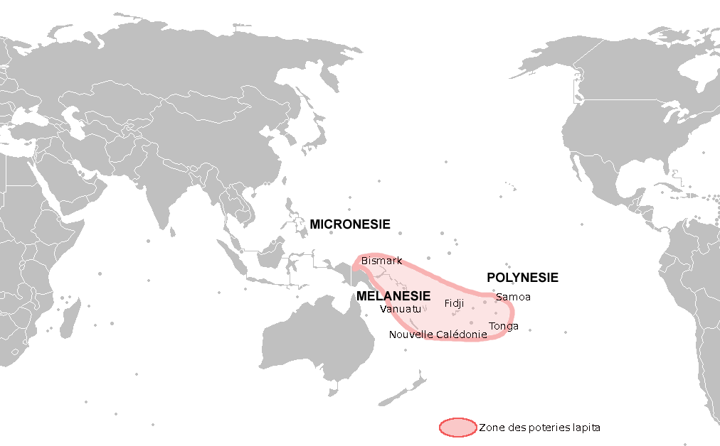

Lapita Expansion
- Easter Island by ca. ACE 300
- Hawai’i by ca. ACE 400
- New Zealand by ca. ACE 1000
Ocean-going Canoes
For more information, see the video at the bottom of the chapter as well as http://www.hokulea.com/

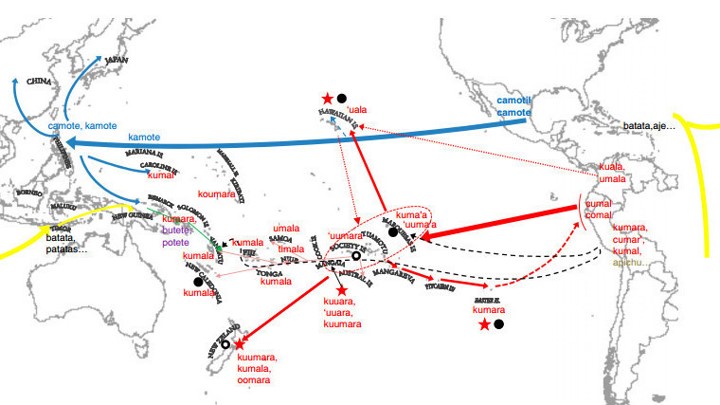
Sweet Potatoes (and Yams)
As Americans, we usually get confused by sweet potatoes and yams as these terms are used interchangeably Also, sweet potatoes are called “yellow yams”! However, they belong to two separate plant families and were domesticated in two, possibly three different regions of the world. The sweet potato is a dicotyledonous (refers to sexual reproduction of seeds with two dicotylous embryonic leaves) plant that belongs to the family Convolvulaceae. Of the approximately 50 genera and more than 1,000 species of Convolvulaceae, Ipomoea batatas is the only crop plant of major importance. Ipomoea batatas is related to the common potato (Solanum tuberosum). There may have been two centers of domestication:
- Lowland Mesoamerica, ca. 3,000 BC
- Tropical South America, ca. 7,000 BC

Key Takeaways
- Distinctive human traits include: bipedalism; large brains; reduced canines; “V” shaped jaw; females with no outward signs of ovulation and sexually receptive throughout the year; high male parental investment; cumulative cultural evolution; and spoken complex language.
- Over the course of human evolution, natural selection favored large brains long after we were habitual bipeds. As a result, human infants are born premature and humans need assistance in birth. Habitual bipedialism has also led to back, knee, and hip problems.
- Throughout our evolution, the tools became more complicated, representing increasing brain size and more complex thought patterns.
- Our cumulative cultural evolution has allowed culture to become the primary means by which we adapt to our environments and allowed our species, Homo sapiens to spread over the entire globe.
- Cooperation between and among members of human social groups was perhaps one of our greatest adaptations.
This refers to projection of the lower facial skeleton that houses dentition beyond some reference point such as the forehead. In primates, humans have the least prognathic face, indeed an orthognathic one, which means essentially vertical.
Recognizing a lack of consensus definition of human or hominin, we define hominin as including all taxa with evidence of bipedalism that have emerged since the split from the common ancestor of humans and chimpanzees/bonobos.
Rifting is the process by which the continental lithosphere gets "stretched;" it breaks apart or becomes separated through deep magma up-welling that pushes tectonic plates apart resulting in faulting and vulcanism.
Foraminifera (forams for short) are single-celled organisms found in all marine environments that have perforated shells commonly composed of calcium carbonate and having distinctive architecture. In death the shells settle to form thick ocean-floor sediments.
A ridge of bone running lengthwise along the middle top of the skull and serving as an anchor for strong muscles to work the jaw. In human evolution the loss of this feature is thought associated with dietary changes leading to the genus Homo.
A space separating teeth of different functions. For primates especially, it is the gap between the incisors (biting teeth) and premolars and molars (grinding teeth) that accommodates large canines. Humans lack this feature.
the whole in the base of the skull where the brain stem passes; can be used to tell locomotive pattern (bipedal or quadrupedal)
This is the simplest and earliest method for fracturing a piece of stone to detached spalls known as flakes that have sharp edges and to produce sharp edges on the rock struck, which is known as the core. The percussion tools used for flaking rocks were originally all made of stone, known as hammerstones. Considerably later in human evolution, some flaking tools started to be made of wood or antler. Such softer tools when used for direct percussion create different types of flakes that are often thinner.
